
Parece imposible creer que cada parte del planeta se encuentre habitada por bacterias en gran número y diversidad. Sin embargo, podemos encontrarlos desde zonas casi inhabitadas como la Antártida hasta en objetos inimaginables como la punta de un alfiler (¡Más de un millón solo en la punta!) Por ello, no es de extrañar que la diversidad del suelo sea tan grande si la comparamos con un alfiler, no solo por su extensión sino, además, por lo diverso y complejo que puede ser este hábitat.
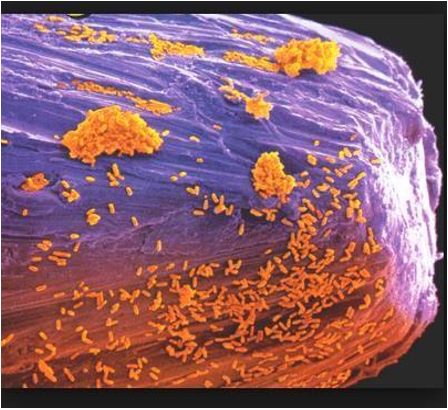
De toda esta extensión, la rizósfera es la zona que presenta mayor actividad biológica y química (Li et al., 2014) y con ello mayor concentración de microorganismos. Se conoce como rizósfera a la zona del suelo que rodea a la ra
piatti di plastica dorati
bershka zenske kozne jakne cijena
כרית פריד קלאסיק
чанти за лаптоп dell
pezzi per pc a basso costo amazon
a little rebellion is a good thing shirt
שואב אבק דייסון v10 דגמין
tiffany and co couple rings
червило в ръчен багаж
закопчалка за колан мерцедес c220 cdi лява и дясна предни
barbie ház fából
παπαγιαννοπουλοσ ταινία ο αχόρταγοσ
filtro askoll pratiko 400
compex quels programme
קמילוטרקט מסכה לשיער 250 מ»
íz de la planta y que comprende no más de 1mm de ancho, desde la zona de la raíz hacia el suelo (Lines-Kelly, 2005), asimismo provee un complejo y dinámico microambiente, donde las bacterias y hongos, en asociación con las raíces, forman comunidades únicas que tienen considerable potencial para la detoxificación de compuestos orgánicos nocivos(Cavallero, 2013). En ella se dan importantes interacciones entre los organismos del suelo y las plantas; tal que muchos compuestos elaborados por estas últimas son transferidos a los microorganismos y viceversa. Como consecuencia, se pueden dar interacciones con efectos tanto positivos como perjudiciales.
En lo que refiere a los efectos positivos (o benéficos) en la zona de la rizósfera, estos son llevados a cabo por un grupo de microorganismos conocidos por sus siglas en inglés PGPR (Plant Growth Promoting Rizobacteria) (Barka et al, 2009). Estos abarcan un conjunto de bacterias que inciden directamente sobre el desarrollo y protección de las plantas distinguiéndose dos subgrupos: PGPR y Biocontrol-PGPR (Kloepper, 1992; Bach y Díaz, 2010; Toro et al, 2013). Los mecanismos llevados a cabo por ellas son diversos y pueden dividirse en estimulación directa y estimulación indirecta:
Estimulación directa:
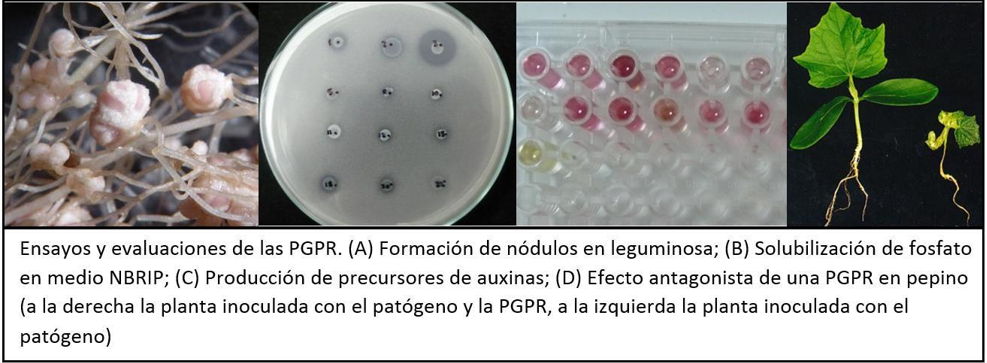
- La fijación de nitrógeno: La capacidad de fijar nitrógeno atmosférico en algunas bacterias es ampliamente conocida, existiendo dos mecanismos por los cuales pueden llevar a cabo el proceso: El primero es a través de la formación de nódulos en leguminosas, los cuales son formados por bacterias del género Rhizobium (Laurette et al, 2015; Jang et al, 2013). La interacción es de tipo mutualista; es decir, plantas y bacterias se benefician mutuamente, mas no es esencial esta asociación para poder existir en el ambiente. En cuanto al segundo proceso, se da a través de la reducción del nitrógeno atmosférico, dando como producto final amonio o nitrato; productos que puede finalmente tomar la planta para el desarrollo de sus estructuras.
- Producción de fitohormonas: La producción de los reguladores de crecimiento se produce al interior de las plantas según su ciclo de desarrollo. Dentro de los principales reguladores se encuentran las auxinas, giberelinas, citoquininas y etileno (Taiz & Zeiger, 2006). Sin embargo, las plantas no son las únicas con la capacidad de producir fitohormonas. Las PGPR tienen pueden sintetizarlas e incluso a sus precursores químicos, con lo cual participan en la regulación del crecimiento y desarrollo de las estructuras de las plantas (Lara et al, 2011; Pandya y Desai, 2014; Raja et al, 2006; Lenin y Jayanthi, 2012; Cassán et al, 2001).
Estimulación indirecta
- Producción de sideróforos: Son estructuras formadas tanto por plantas como por bacterias que permiten secuestrar el hierro (Fe) de las sales a las que se encuentra unido, haciéndolo biodisponible para ambos (Aguado- Santacruz et al, 2012; Katiyar y Goel, 2003). Existen estudios que afirman que estas estructuras participan, además, en el control de fitopatógenos, evitando la toma de los metales por los patógenos (competencia por nutrientes) (Martínez-Viveros et al., 2010).
- Efectos antagónicos e inducción de resistencia a patógenos: Muchos microorganismos tienen la capacidad de producir una gran variedad de compuestos que resultan perjudiciales para otros (antibióticos) los cuales juegan un rol importante en el control biológico (Raaijmakers et al., 2002). Ese es el caso de muchas PGPR, las cuales producen compuestos antagónicos de diferente naturaleza y estructura. Todos ellos, producidos para eliminar competidores de la rizósfera, que en muchos casos resultan ser patógenos de plantas (Barka et al, 2000). Dentro de este mecanismo de control: la producción de enzimas hidrolíticas, la competencia por nutrientes o nicho ecológico en la raíz y la regulación de la producción de etileno son los más conocidos dentro del conjunto de estrategias que poseen para asegurar su predominancia en la zona (Compant et al., 2005). La inducción del sistema de resistencia de las plantas por las PGPR es otra forma de asegurar la colonización y protección de la planta tanto en la rizósfera como en toda la planta, mas no se considera un efecto antagonista sino una característica de agente bio-controlador (Beneduzi et al, 2012). La estimulación del sistema por compuestos producidos por las bacterias genera la producción de sustancias en la planta que finalmente la protegerán de los agentes externos (Zdor y Anderson; 1992).
- Solubilización de fósforo: Dentro de todos los elementos, el fósforo es uno de los tres elementos más importantes dentro de la nutrición de las plantas. Sin embargo, este se encuentra en baja disponibilidad dentro del suelo; ya que según el tipo de suelo puede encontrarse asociado al hierro o al aluminio (en suelos ácidos) o al calcio (en suelos neutros o alcalinos) y en otros casos puede encontrarse en complejos orgánicos (Mackenzie & Amer, 1964; Confort, s/f). Para hacer uso del P, los microorganismos despliegan diversos mecanismos de solubilización: producción de ácidos orgánicos e inorgánicos; vía enzimática a través de fosfatasas y como mecanismo principal, la secreción de protones (Behera et al, 2013). En lo que refiere a los ácidos orgánicos, estos bloquean los sitios de absorción del P o forman complejos con los cationes en la superficie mineral del suelo (Khan et al, 2009). En cuanto a la vía enzimática las fosfatasas pueden ser de dos tipos: alcalina o ácida. A su vez, dentro de las fosfatasas ácidas encontramos un grupo particular por su capacidad de solubilizar fitatos (fósforo orgánico), las fitasas (Behera, 2013). Sea cual sea el mecanismo empleado por la bacteria, su uso incrementa la captación de P por las plantas. Ello permite el uso de diferentes aditivos como fuente de P tales como la roca fosfórica, hidroxiapatita, entre otros.
Usos en la agricultura
El uso de las PGPR inicia muchos años atrás, antes de la elaboración de productos formulados a base de microorganismos benéficos del suelo, inicia con la mezcla de semillas y suelo inoculado por procesos naturales (Bach & Díaz, 2008). Años más tarde se elaborarían los primeros inoculantes (y patentes) de productos a base de cepas de Rhizobium sp, Bacillus sp, Azotobacter sp., Pseudomonas putida, Pseduomonas fluorescens, entre otros (Bach & Díaz, 2008).
Dentro de las formulaciones de inoculantes bacterianos podemos encontrar dos tipos de productos biológicos: los que se emplean en el control de enfermedades y los que se emplean para la estimulación del crecimiento de las plantas (Bach & Díaz, 2008). Estos no deben ser confundidos con los productos biológicos formulados, los cuales son otro tipo de bioinsumos que pueden estar constituidos por extractos vegetales, fitohormonas o los metabolitos bacterianos (SENASA, s/f).
Ideas finales
En la actualidad, en el Perú existe una variedad de productos a base de uno o más microorganismos dentro de la formulación. Sin embargo, a pesar de la variedad de productos, el mercado aún es incipiente; esto debido a que a la fecha aún su difusión es baja. Por otra parte, varios de los productos distribuidos no se encuentran regulados debidamente en lo que refiere al control de calidad, con lo cual no existe un cuidado hacia la conservación del producto, por ello es importante asegurarnos de la confiabilidad hacia el proveedor y la de sus procesos. Esto último supone una desventaja para el agricultor tras adquirir el producto, ya que muchas veces las malas prácticas de las empresas distribuidoras (mal almacenamiento, venta de productos vencidos, distribución en condiciones ambientales desfavorables, entre otros) hacen que el producto pierda efectividad hasta casi en un 100%. En consecuencia, la desconfianza a productos alternativos a los fertilizantes químicos pierde credibilidad y confianza haciendo su establecimiento más difícil.
El cambio climático es un efecto innegable, el cual nos exige una mirada a las alternativas biotecnológicas que dentro de unos años serán la clave para un futuro sustentable y una productividad continua. Por ello es importante tomar conciencia de la importancia que lleva el uso de productos biotecnológicos debidamente regulados para el cuidado del medio ambiente.
Incluso, los estudios de los potenciales microorganismos PGPR’s en conjunto con otros productos de uso agrícola como el compost corroboran la posibilidad de obtener mayores rendimientos en las campañas, siendo su uso ampliamente respaldado por más de un estudio efectuado en cultivos de gran importancia como son la papa (El-Sayed et al., 2014), maíz (Saber et al., 2012), tomate (De Brito et al., 1995; Haggag & Saber, 2000), entre otros.
Como se ve, actualmente, las nuevas tendencias y tecnologías nos llevan a optar por el uso de alternativas que vayan de la mano del cuidado del medio ambiente. Por ello, tomar la alternativa del uso de las PGPR de la mano de productos que actúen como suministros de nutrientes (como lo es el compost), son la clave para una continuidad en la actividad agrícola.
Nosotros, FOSAC, como empresa responsable con el medio ambiente y su entorno, estamos comprometidos con el cuidado y conservación de los suelos del Perú, por ello contamos con un producto acorde a las necesidades nutricionales y microbiológicas de los mismos. MO- STD Aqua, es un mejorador de las propiedades físicas, químicas y biológicas del suelo que proporciona además de una mezcla de probióticos y prebióticos activos, los ácidos húmicos , que es finalmente la fracción activa de la materia orgánica, desarrollando microorganismo benéficos que poseen actividad antagónica contra: Verticilium, Fusarium, Phytophthora y nematodos, entre otros patógenos.
Bióloga Alexandra Florián Silva
Bibliografía
Aguado-Santacruz, G.; Moreno-Gómez, B.; Jiménez-Francisco, B.; García-Moya, E. y Preciado- Ortiz, R. 2012. Impacto de los sideróforos microbianos y fitosideróforos en la asimilación de hierro por las plantas: una síntesis. Revista Fitotecnia Mexicana 35(1):9-21
Bach, T.; Díaz, M. 2008. Las Rizobacterias Promotoras de Crecimiento Vegetal (PGPR) en la agricultura. Agricultura orgánica. 3:35-38
Barka, E.; Belarbi, A.; Hachet, C.; Nowak, J. y Audran, J. 2000. Enhancement of in vitro growth and resistance to gray mould of Vitis vinifera co-cultured with plant growth-promoting rhizobacteria. FEMS Microbiology Letters. 186:91-95
Behera, B.C.; Singdevsachan, S.; Mishra, R.; Dutta, S. y Thatoi, H. 2013. Diversity, mechanism and biotechnology of phosphate solubilising microorganism inmangrove—A review. Biocatalysis and Agricultural Biotechnology. 3(2):97-110
Beneduzi A., Ambrosini, A. y Passaglia, L. 2012. Plant growth-promoting rhizobacteria (PGPR): Their potential as antagonists and biocontrol agents. Genetics and Molecular Biology. 35(4):1044- 1051
Cassán, F.; Bottini, R. y Piccoli, P. 2001. In Vivo Gibberellin A Metabolism by Azospirillum sp. in dyDWARF rice mutant seedlings. Proc. Plant Growth Reg. 28:124-129.
Cavallero, G. J. 2013. Prefacio. Tesina- Caracterización de Aislamientos de Microorganismos Nativos Simbiontes de Rhynchosia, 1–61.
Compant, S.; Duffy, B.; Nowak, J.; Clément, C. y Barka, E. 2005. Use of Plant Growth-Promoting Bacteria for Biocontrol of Plant Diseases: Principles, Mechanisms of Action, and Future Prospects. Applied and Environmental Microbiology. 71(9): 4951-4959.
Compant, S.; Duffy, B.; Nowak, J.; Clément, C. y Barka, E. 2005. Use of Plant Growth-Promoting
Cornforth, I. The Fate of Phosphate Fertilisers in Soil. Chemicals and Soils-D-Phosphate. En línea: http://nzic.org.nz/ChemProcesses/soils/2D.pdf
De Brito, M.; Gagné, S. y Antoun, H. 1995. Effect of Compost on Rhizosphere Microflora of the Tomato and on the Incidence of Plant Growth-Promoting Rhizobacteria. Applied and Environmental Microbiology. 61(1):194-199
El-Sayed, S.; Hassan, H.; El-Mogy, M. y Abdel-Wahab, A. 2014. American-Eurasian Journal Agriculture and Environment Science. 14(7): 636-643.
Haggag, W. & Saber, M. 2000. Use of compost formulations fortified with plant growth promoting rhizobacteria to control root-rot diseases in some vegetables grown in plastic houses. En línea: http://www.acgssr.org/BioTechnology/V4N1January2001/fullpaper/p08.PDF
Jang, J.; Kwon, M.; Park, D.; Sung, C. y Kim, C. 2013. Screening of Rhizobium, Hairy Vetch Root Nodule Bacteria, with Promotion of Nodulating and Nitrogen Fixation. Korea Agricultural Science Digital Library. 49(2):131-136
Katiyar, V. y Goel, R. 2004. Siderophore mediated plant growth promotion at low temperature by mutant of fluorescent pseudomonad. Plant Growth Regulation 42:239-244.
Khan, A.; Jilani, G.; Akhtar, M.; Saqlan, S.; Rasheed, M. 2009. Phosphorus Solubilizing Bacteria: Ocurrence, Mechanisms and their Role in Crop Production. J. Agri. Biol. Sci. 1(1):48-58.
Kloepper, J. 1992. Plant growth-promoting rhizobacteria as biological control agents. In FB Metting Jr, ed, Soil Microbial Ecology: Applications in Agricultural and Environmental Management. Marcel Dekker Inc., New York, pp 255-274
Lara, C.; Oviedo, L. y Betancur, C. 2011. Bacterias nativas con potencial en la producción de ácido indolacético para mejorar los pastos. Zootecnia Trop. 29(2):187-194.
Laurette, N.; Bisseck, N.; Henri, F.; Souleymanou, A.; Kandem, K.; Albert, N.; Dieudonné, N. y Francois, E. 2015. Isolation and Screening of Indigenous Bambara Groundnut (Vigna Subterranea) Nodulating Bacteria for their Tolerance to Some Environmental Stresses. American Journal of Microbiological Research. 3(2): 65-75
Lenin, G. & Jayanthi, M. 2012. Indole Acetic Acid, Gibberellic Acid and Siderophore Production by PGPR Isolates from Rhizospheric Soils of Catharanthus roseus. International Journal of Pharmaceutical & Biological Archives. 3(4): 933-938.
Li, X.; Rui, J.; Xiong, J.; Li, J.; He, Z.; Zhou, J.; Yannarell, A. y Mackie, R. 2014. Functional Potencial of Soil Microbial Communities in the Maize Rhizosphere. PLoS ONE 9(11):1-9
Lines-Kelly, R. 2005. The Rhizosphere. Soil biology basics. En línea: http://www.dpi.nsw.gov.au/ data/assets/pdf_file/0004/42259/Rhizosphere.pdf
Mackenzie, A.; Amer, S. 1964. Reactions of iron, aluminum and calcium phosphates in six Ontario soils. Plant and Soil XXI. 1:17-25
Martínez, O.; Jorquera, M.; Crowley, D.; Gajardo, G. y Mora, M. 2010. Mechanisms and Practical Considerations Involved in Plant Growth Promotion by Rhizobacteria. J. Soil Science Nutr. 10(3):293-319.
Pandya, N. y Desai, P. 2014. Screening and characterization of GA3 producing Pseudomonas monteilii and its impact on plant growth promotion. Int. J. Curr. Microbiol. App. Sci. 3(5):110-115
